
Die Asteridae sind die evolutionär am höchsten stehenden Angiospermen. Sie erschienen kurz vor Beginn des Tertiärs und entfalteten sich bereits während des Oligozäns sehr stark. Sie zeichnen sich durch Sympetalie aus, d.h., ihre Petalen sind zu einer Kronröhre verwachsen. Die Zahl der Stamina ist gleich oder geringer als die der Kronröhrenzipfel. Pflanzen mit verwachsenen Petalen wurden von A. ENGLER (1887) zur Gruppe der Sympetalae zusammengefaßt. Sie ist mit der der Asteridae nahezu deckungsgleich, wenngleich von ihnen einige nicht besonders artenreiche Ordnungen abgetrennt werden mußten: Diapensiales, Ericales, Ebenales, Primulales, Plumbaginales und Cucurbitales.
Die Blüten der Asteridae enthalten Nektarscheiben, und man vermutet, daß diese dem inneren Staubblattkreis homolog seien; sie sind nie polyandrisch. Die Stamina sitzen direkt an der Corolla. Der Fruchtknoten besteht aus meist zwei miteinander verwachsenen Karpellen. Alle Arten der Placentation kommen vor. Der Pollen ist triaperturat (meist tricolpat). Die Samen sind in der Regel endospermhaltig und die Embryonen zweikeimblättrig; lediglich bei einigen Parasiten und Arten mit mycotropher Lebensweise (obligate Symbiose mit Pilzen) entfällt die Differenzierung in die beiden Keimblätter.
Die Asteridae verfügen über ein weites Spektrum an sekundären Pflanzenstoffen. Die Substanzen sind durchweg Produkte verkürzter Biosynthesewege, d.h., ihre Produktion ist weniger energieaufwendig als die von Produkten primitiverer Pflanzengruppen. Krautige Pflanzen sind unter den Asteridae vorherrschend. Infolge ihrer kürzeren Generationsdauer ist bei ihnen die Chance, neue Genkombinationen zu erproben, höher; damit auch die Chance, neue Lebensräume zu besiedeln. Diese Strategie ist an einer Erhöhung der Artenzahl (im Vergleich zur Artenzahl der Holzpflanzen) erkennbar. Vorhanden sind verschiedene Alkaloide (aber niemals Benzylisochinolinalkaloide), dann Polyacetylene und iridoide Verbindungen , während cyanogene Verbindungen und Saponine relativ selten sind, Ellagitannine und kondensierte Gerbstoffe nur bei sehr wenigen Arten vorkommen, und Betalaine und Senföle nie nachgewiesen wurden. Das Fehlen der Betalaine und Isochinolinalkaloide kann zur Abgrenzung der Asteridae gegenüber den Caryophyllidae und den Magnoliidae benutzt werden. Das nur ausnahmsweise Vorkommen der Ellagsäure und der Proanthocyanine trennt sie von den Rosidae, Dilleniidae und Hamamelididae. Diese Aussage ist nicht "wasserdicht", denn einer großen Zahl von Arten der drei zuletzt genannten Subklassen fehlen diese Substanzen ebenfalls. Die in einigen Familien (Loganiaceae, Apocynaceae und Rubiaceae) vorkommenden Indolalkaloide sind die giftigsten Verbindungen dieser Art überhaupt. Sie entstanden aus der Koppelung zweier weit verbreiteter Verbindungen: den Iridoiden, die aus Monoterpenen entstehen, und dem Indolring, der aus Trytophan hervorgeht.
Es gibt gute Argumente für die Annahme, daß die Asteridae von Vorfahren der Rosales abstammen, denn alle Merkmale, die für die Asteridae typisch sind, findet man getrennt auch bei einzelnen Arten, Gattungen oder Familien der Rosales. Der evolutionäre Fortschritt der Asteridae wäre demnach in einer Kombination bereits vorhandener Merkmale (bzw. deren Erbanlagen) zu neuen funktionellen Einheiten zu sehen. Es sei vermerkt, daß ihre Verwandtschaft zu den meisten rezenten Ordnungen der Rosidae allerdings kaum erkennbar ist, weil jene bereits zu spezialisiert sind. Die Blüten der Asteridae sind in vieler Hinsicht an die Lebensweise und das Mustererkennungsvermögen ihrer Bestäuber angepaßt. Auffallend ist eine Tendenz zur Zygomorphie. Während radiärsymmetrische Blüten in ihrer Größe beträchtlich variieren, ist die Variation der Größe zygomorpher Blüten stark eingeschränkt und an die Körpergrößen ihrer Bestäuber adaptiert. Das Vorhandensein von Landeplätzen, Nektarien, Blütenmalen und die Produktion von Duftstoffen sind weitere Spezialisierungen, die der Anpassung an Bestäuber dienen.
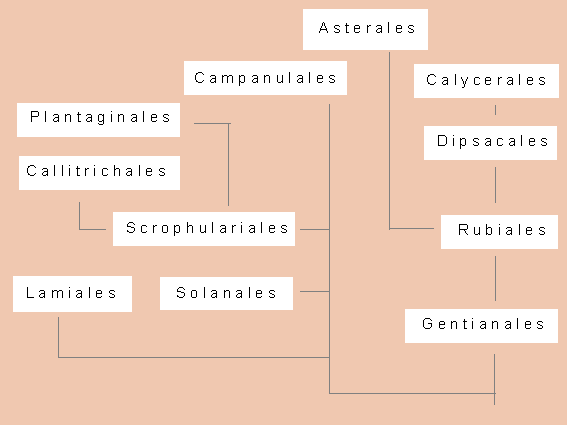
Asteridae: Ein phylogenetisches Schema zur Darstellung der Beziehungen zwischen den einzelnen Ordnungen (Nach A. CRONQUIST, 1981)
Den Asteridae gehören 60 000 Arten in 11 Ordnungen und 49 Familien an. Wie bereits dargelegt, weist die geringe Zahl an Ordnungen auf eine weitgehende Homogenität dieser Unterklasse hin. Offensichtlich wurde auf der Ebene der Rosidae und der Parallelgruppe der Dilleniidae alles das ausprobiert, was das Angiospermengenom durch Umstrukturierung hergibt. G. L. STEBBINS (1984) nennt diesen Prozeß eine Mosaikevolution. Auf der Ebene der Asteridae wurde eine optimale Kombination gefunden, und es ging im Verlauf ihrer Evolution vornehmlich um eine Optimierung von Details im Blütenbau. Die Wechselwirkungen und gegenseitigen Abhängigkeiten von Blüten und Bestäubern (meist Insekten) wurden vervollkommnet. Eine solche Anpassung setzt aber auch voraus, daß die Evolution der Insekten mit der der Blüten Schritt hielt.
Ein alternativer Klassifikationsvorschlag:
Wie bereits erwähnt, sind Alternativen zu einem System der Kategoriezuordnungen möglich - sofern sachlich begründbar -, so untergliedern D. FROHNE und U. JENSEN (1985) die Asteridae (s. l.) in die Cornidae, wobei hier einige Ordnungen zugeschlagen werden, die bereits bei den Rosidae behandelt wurden, die Lamiidae und die Asteridae (s. str.). Diese Zuordnung beruht auf der Gruppierung der folgenden Merkmalskombinationen:
Cornidae: Blätter einfach, ungeteilt; oft sympetale Blüten mit coenokarpem Gynaeceum; scalariforme Perforation in Gefäßen. Das ist ein primitives Merkmal und ein Hinweis darauf, daß die Cornidae den Hamamelididae und den Magnoliidae nahestehen. Das Endosperm ist zellulär ausgebildet. Iridoide sind die vorherrschenden sekundären Pflanzenstoffe. Folgende Ordnungen werden zu dieser Unterklasse zusammengefaßt: Cornales, Dipsacales, Sarraceniales, Ericales, Gentianales, Oleales (hier als eigenständige Ordnung geführt).
Lamiidae: Es sind, wie die Cornidae, Iridoidpflanzen, was dafür spricht, daß beide Taxa phylogenetisch zusammengehören. Polyacetylene fehlen. Man findet bei ihnen im Vergleich zu den Cornidae mehr abgeleitete Merkmale und Merkmalskombinationen. Vorwiegend Kräuter. Die wechsel- oder gegenständigen Blätter tragen keine Nebenblätter. Die Blüten sind tetrazyklisch d.h., sie verfügen über nur einen Staubblattkreis. Die Filamente der Staubblätter sind an der Basis mit der Kronröhre verwachsen. Zygomorphe Blüten besitzen vier, radiärsymmetrische fünf Staubblätter (in Einzelfällen ist die Zahl sogar noch niedriger). Es sind nur zwei Fruchtblätter vorhanden. Zugehörige Ordnungen: Solanales, Boraginales (hier als eigenständige Ordnung geführt), Scrophulariales, Lamiales.
Asteridae (s.str.): Keine Iridoidpflanzen, sie enthalten statt dessen Polyacetylene als sekundäre Pflanzenstoffe. Die meist fünf Blütenblätter sind hier zu einer Kronröhre verwachsen, zwei Fruchtblätter, meist fünf Staubblätter (nie polyandrisch). Der Pollen ist trinukleat. Es bestehen Tendenzen zur Bildung dorsiventraler Blüten und einer Verwachsung von Antheren. Nebenblätter fehlen. Zugehörige Ordnungen sind Campanulales und Asterales.
Derzeit in Botanik online nicht besprochene Odnungen: Callitrichales, Calycerales: Familien und Gattungen
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de