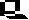
|
|
|
|
|
EVOLUTION |
|
Population genetics This statement of the Hardy Weinberg Law describes a state of equilibrium in the genetic makeup of populations that is more interesting for its exceptions than the comparitively rare occasions when it occurs. Looking at the gene frequencies in the dihybrid cross we see that the frequency of dominant and recessive alleles remains constant at 50% from P1(parental) to F1 and F2 generations. Even if we were to select for the phenotype of the dominant genes, recessive alleles would persist in the population for several generations because they would be concealed by the dominant alleles in the heterozygous state. When recessives occur at low frequency the chances of them coming together in the homozygous state become very low, so that even a lethal recessive can persist for many generations. Exceptions to the rule lead to genetic change, or evolution in populations:
So it is doubtful that the Hardy Weinberg equilibrium exists for long periods of time in real-life populations. Even in the absence of direct selection, organisms can be subject to"genetic drift". Problems of genetic uniformity Similar considerations apply to human manipulation of crop plants. The production of uniform stands of crops with consistent composition and quality is attractive but brings with it the risk of uniform susceptibility to a particular disease. A famous example is the problem of early hybrid corn varieties that were all susceptible to a new race of Southern corn blight (Helminthosporium maydis) that arose in 1970. About 15% of the US corn crop was lost. Selection Human beings have become major agents of selection, both conscious and unconscious. Many of our crop plants are far-removed from their wild ancestors so that it can even be difficult to say where they came from. Corn (Zea mays) is so unlike any wild grass that we are still trying to explain its origin. An example of unwitting plant selection is the development of metal tolerance in bent grass (Agrostis tenuis) in the Upper Swansea Valley. This is an area where soils became heavily polluted with copper and other metals in the nineteenth century. Bent grass from nearby unpolluted areas is unable to colonize these soils, but the species is found growing there. In the laboratory the grass from polluted areas can be shown to be unusually tolerant to copper. It does not persist when transplanted to unpolluted areas where it cannot compete with non-tolerant strains. Evolutionary divergence |
|
|
|
|
|
|
|
|
In mountainous parts of the US, such as California the environment changes substantially over short distances. Some plants, such as sticky cinquefoil (Potentilla glandulosa) occur from sea level up to 5,000 feet. There are distinct ecotypes that are adapted to different altitudes. A low altitude form transplanted to the mountains fails to make enough vegetative growth in the short summer season and dies in the following winter. When plants colonize a new environment, such as an island, with little existing vegetation they have an unusual opportunity to develop a variety of forms to fill different ecological niches. This leads to adaptive radiation, exemplified particularly well in plants like the tarweeds or silverswords (Dabautia, Wilkesia, and Argyroxiphium) on Hawaii and other Pacific islands. Hybridization
The development of mechanisms to encourage or ensure outbreeding can be understood in relation to the advantages of heterozygosity. Deliberate crossing of inbred lines of crop plants often leads to an increase in plant growth and yield (Hybrid vigor or heterosis). This is exploited in the production of F1 hybrids of crop and ornamental plants. Hybrid vigor may be particularly pronounced when different species or even genera are crossed. The London plane (Platanus x acerifolia) is a cold-tolerant urban tree, produced by crossing the North American sycamore (Platanus occidentalis) with the tender oriental plane (Platanus orientalis). |
|
|
The London plane is fully fertile but many interspecific hybrids are sterile. Although the genomes of the parental species are related, differences in chromosome structure prevent normal meiotic paring and separation so that functional spores are not formed. Hybrids of annual plants would inevitably die because of this, but hybrids of perennial plants can persist and even spread vegetatively. An example is the common horsetail. Equisetum x ferrisii is a sterile hybrid of E. laevigatum and E. hyemale. The hybrid has spread along rivers in the midwest because pieces of rhizome get carried along on the stream. |
|
|
Fertility can be restored by doubling the number of chromosomes; then every chromosome can find an exact match with which to pair during meiosis. This can happen by chance in nature as in the example of the grass Spartina anglica a tetraploid derivative of a hybrid between a North American species (S. alterniflora) and the European S. maritima. Spartina anglica has colonized large areas of coastal Britain since its origin at the turn of the century. |
|
|
Many crop plants are allopolyploids originating from duplication of the genomes of interspecific hybrids. Examples of allotetraploids among the Brassica vegetables have already been discussed in lab. Wheat, Triticum aestivum and strawberry, Fragaria ananassa are examples of allohexaploids. More recently the deliberate induction of polyploidy by colchicine treatment has become a standard practice in plant breeding in order to obtain fertile hybrids. Hybrid vigor is stabilized in a plant with two or more genomes between which there is no recombination. Polyploids are also more vigorous than their diploid ancestors, even if there is no mixing of genomes (autopolyploids contain multiple sets of a single haploid genome.) |
|
|
|
|
|
Another escape from sterility in hybrid plants is apomixis, the production of an embryo from a diploid cell without meiosis and gamete fusion. This occurs in many hawthorns and brambles and has the same advantages and disadvantages as vegetative reproduction. Seed production is assured and a successful genotype can rapidly invade a suitable habitat but there is little prospect of future variation to adapt to changing circumstances unless the sexual process can be restored.
Copyright © Michael Knee, |
|