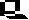

In den beiden vergangenen Jahrzehnten sind eine Reihe von Hochrechnungen durchgeführt worden, die unter Verwendung vorhandener Meßwerte das Ziel verfolgten, zukünftige Entwicklungen vorherzusagen, um Politikern Entscheidungshilfen bei der langfristigen Planung zu bieten. Besonders intensiv befaßte sich der "Club of Rome", eine Vereinigung von Wissenschaftlern mit Interesse an Zukunftsfragen, mit der Problematik des globalen Gleichgewichts sowie mit Modellstudien zur Wachstumskrise der menschlYchen Bevölkerung und den Auswirkungen der Umweltzerstörung. Zahlreiche Simulationen wurden während der siebziger Jahre in den Arbeitsgruppen von D. L. MEADOWS am Massachussetts Institute of Technology und E. PESTEL, Technische Universität Hannover, angestellt. Sämtliche Kalkulationen beruhten auf folgenden Prämissen:
- Die Zuwachsraten bleiben unverändert. Die Systemvariablen sind voneinander abhängig.
- Eine der Variablen wird rechnerisch manipuliert, um das Verhalten der übrigen Parameter im System zu studieren.
Alle Annahmen gehen davon aus, daß die Ressourcen der Erde endlich sind und daß irgendeine Veränderung in dem derzeitigen exponentiellen Wachstumsprozeß (Bevölkerungsexplosion, Umweltzerstörung u.a.) notwendig sein wird. Zu den endlichen Ressourcen zählen:
- Der Vorrat an ausbeutbaren, nicht erneuerbaren Rohstoffen.
- Die Absorptionskapazität der Umwelt für Schadstoffe.
- Die Reserven an potentiellem Ackerland.
- Der Ertrag pro Hektar Land.
Niemand kennt die Grenzen genau. Auch sind sie nur schwer durch jeweils eine einzige Zahl zu definieren, da diese sich durch Mißbrauch von Ressourcen (überhöhter Energieverbrauch in Industrienationen, Raubbau an der Natur in Entwicklungsländern und vieles mehr) reduziert.
Bei jeder Modellkonstruktion sind Rückkopplungsschleifen von eminenter Bedeutung. Sie beschreiben Vorgänge, bei denen das Endprodukt auf das Ausgangsprodukt wirkt. Dabei müssen Verzögerungsfaktoren berücksichtigt werden, denn viele Auswirkungen treten erst mit einer zeitlichen Verzögerung auf, was zur Folge hat, daß manche Entwicklungen durch ein "Überschwingen" gekennzeichnet sind. Geregelte Systeme sind durch eine negative Rückkopplung charakterisiert, denn das Endprodukt hemmt seine Eigensynthese. Durch eine positive Rückkopplung ("Wachstum fördert Wachstum", siehe Wachstumsfunktion ) kann ein System eskalieren und aus dem Gleichgewicht geraten. Von den zahlreichen veröffentlichten Ansätzen wurde zur folgenden Darstellung der Modellfall "Eutrophierung von Seen" herausgegriffen (J. M. ANDERSON, 1973). Doch zuvor einige Begriffe und Definitionen ( s. a. Eutrophierung von Gewässern):
Oligotropher See (oligotrophes Gewässer, Oligotrophie): nährstoffarmer See, Nährstoffarmut. Beispiele: Gebirgsseen und viele Seen in Skandinavien.
Eutropher See (eutrophes Gewässer, Eutrophie): nährstoffreicher See, Nährstoffreichtum. In diese Kategorie fallen u.a. die mitteleuropäischen (stehenden) Gewässer. Unter Eutrophierung versteht man den Prozeß zunehmender Nährstoffzufuhr und damit einer Nährstoffanreicherung in Gewässern. Zu den am stärksten belastenden Nährstoffen gehören: Phosphate, Nitrate und andere anorganische Substanzen sowie organische Materialien (ungeklärte Abwässer u.a.).
Detritus: partikuläre, amorphe tote organische Substanz (z.B. im Faulschlamm oder im Wasser schwebend).
Epilimnion: obere, lichtdurchstrahlte und damit erwärmte Wasserschicht in einem stehenden Gewässer.
Hypolimnion: untere, unbelichtete und damit kalte Wasserschicht in stehenden Gewässern. Zwischen Epi- und Hypolimnion bildet sich eine Temperatursprungschicht aus (Metalimnion). In einer Zone von weniger als einem Meter Mächtigkeit kann der Temperaturabfall bis zu 10 Grad C betragen.
Die klare Trennung zwischen Epi- und Hypolimnion ist bei Seen in Klimazonen mit Jahreswechsel nur im Sommer ausgeprägt. Während der Wintermonate kommt es zu einer Durchmischung des Wasserkörpers und damit zur Aufhebung des Temperatursprungs.
Die Modellrechnung beruht auf der Annahme, daß Seen durch eine kontinuierliche Phosphatzufuhr eutroph werden. Ein typisches Beispiel hierfür ist der Bodensee, der noch bis ins 20. Jahrhundert hinein oligotroph war. Inzwischen ist die Eutrophierung lokal weit fortgeschritten, 1960 betrug der Phosphatgehalt 20 mg / Kubikmeter, und die Planktonproduktion nahm im Verlauf der letzten 100 Jahre auf das 20fache zu. Die Ursachen für die erhöhte Phosphatzufuhr waren:
- Überdüngung von landwirtschaftlichen Flächen mit Phosphaten (diese Aussage gilt entsprechend auch für Nitrate u.a.).
- Phosphate - als Bestandteile von Waschmitteln (Weichmachern) - gelangen als Abwasser in Flüsse und Seen. In nur wenigen Kläranlagen wird der Phosphatgehalt reduziert (technisch ist dies möglich).
Mit steigender Eutrophierung nimmt der Bestand an Lebewesen zu (gemessen nach Arten- und Individuenzahlen), mit ihnen steigt aber auch der Bedarf an wassergelöstem Sauerstoff.
Vereinfacht dargestellt, ergibt sich die folgende Kausalkette: Nährstoffzufuhr führt zu erhöhter Algenproduktion im See. Es wird daher zunächst zusätzlich Sauerstoff freigesetzt. Da photosyntheseaktive Arten in großer Menge nur im Epilimnion auftreten, dazu auch nur nahe der Seeoberfläche und es in stehenden Gewässern im Sommer kaum eine Wasserumwälzung gibt, geht ein hoher Anteil des gebildeten Sauerstoffs direkt an die Atmosphäre verloren. Nach dem Absterben der Algen erfolgt der stark sauerstoffzehrende bakterielle Abbau, die Sauerstoffkonzentration des Sees sinkt drastisch ab. Stark sauerstoffbedürftige Arten sterben als erste. Am Boden des Sees sammelt sich Detritus (eine Faulschlammschicht). Die Fäulnisprozesse verbrauchen die verbliebenen Sauersstoffreste, und als Stoffwechselprodukte entstehen toxische Kohlenwasserstoffe, so zum Beispiel das Methan u.a. Dies wiederum führt zu einer weiteren Beeinträchtigung des Lebens im See. Das Endstadium ist das sogenannte "Umkippen"; der See ist damit biologisch tot. Als Extrembeispiel für einen solchen Degenerationsprozeß wäre der Erie-See im Norden der USA zu nennen.
Oligotrophe Seen sind in der Regel sauerstoffgesättigt. In eutrophen Seen fällt der Sauerstoffgehalt auf ca. 30 Prozent des Sättigungswerts (vor allem im Hypolimnion). Eutrophierung an sich bedeutet noch lange nicht Absterben allen Lebens im See. Die Mehrzahl eutropher Seen (u.a. fast alle mitteleuropäischen) befinden sich in einem biologischen Gleichgewicht. Eutrophierung ist nämlich ein Prozeß, der in fast jedem See, auch in einer vom Menschen unbelasteten Umwelt in Erscheinung tritt. Natürliche Eutrophierung spielt sich aber in Zeiträumen von Tausenden von Jahren ab und ist ein erster Schritt einer Sukzession:
oligotropher See > eutropher See > Verlandung > Niederungsmoor > Hochmoor
Demgegenüber läuft Eutrophierung durch anthropogene Einflüsse in nur wenigen Jahrzehnten ab; in einem solchen Zeitraum ist an eine Sukzession nicht zu denken. Es lassen sich nunmehr Maßnahmen aufzählen, die einer Eutrophierung entgegenwirken könnten:
Gründliche Behandlung der Abwässer mit dem Ziel, Nährstoffe zu entfernen.
Weitläufige Verteilung von Abwässern. Einleitung in schnellfließende Flüsse (Vorsicht: deren Kapazität und Regenerationsvermögen ist zwar höher als das stehender Gewässer, aber Kapazitätsgrenzen gibt es auch dort).
- Zugabe von Algiziden.
- Entfernung von Detritus vom Gewässerboden durch Ausbaggerung.
Abernten von Algen (Vorteil: sie können getrocknet als Viehfutter verwendet werden).
- Sauerstoffzufuhr durch künstliche Belüftung.
Unter Berücksichtigung dieser
Vorgaben läßt sich ein Sauerstoff- und Nährstoffkreislauf
formulieren. Um die Reaktion des Modells zu testen, benötigt
man quantitative Angaben für die einzelnen Parameter. Als
solche können folgende Werte genommen
werden. Es sind Durchschnittswerte, die in letzter Zeit an einer
Anzahl eutroph werdender Seen gemessen wurden und hier als Eckdaten
für die nachfolgende Modellrechnung herangezogen werden sollen.
Die zu einem Blockschaltbild (Flußdiagramm) zusammengefaßten
Angaben bilden ihrerseits die Grundlage für ein Computerprogramm.
Mit Hilfe des Rechners lassen sich dann unter gegebenen Voraussetzungen folgende Ergebnisse erzielen:
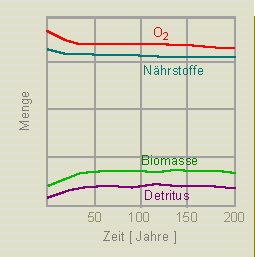
1. Verlauf einer Eutrophierung unter natürlichen Bedingungen. Als Voraussetzung wird eine Nährstoffzufuhr von 1 mg/l/Jahr angegeben. Das System befindet sich im Gleichgewicht. Der Gehalt an Biomasse (und Detritus) nimmt nur unmerklich zu, der Sauerstoffgehalt bleibt annähernd gleich. Unter der Annahme einer Nährstoffzufuhr von 10 mg/l/Jahr ändert sich an der Situation nur wenig.

2. Steigerung der Nährstoffzufuhr. In diesem Fall beginnt man wieder mit einer Nährstoffzufuhr von 10 mg/l/Jahr, steigert den Wert aber jährlich um 2 Prozent. Das genügt, wie die Abbildung zeigt, zum Systemzusammenbruch - der See kippt um.
Erste Hilfsmaßnahme ist die Anwendung von Algiziden: Als Folge dieser Maßnahme entsteht ein sauerstoff- und nährstoffgesättigter See. Gleichzeitig nehmen Biomasse und Detritus den Wert 0 an. Wir erhalten ein totes Gewässer.
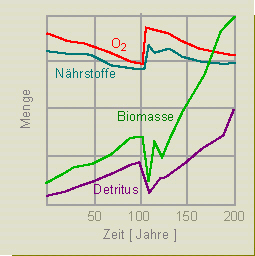
3. Zweite Hilfsmaßnahme: Sauerstoffzufuhr durch künstliche Belüftung. Kurzzeitig führt die Maßnahme zu Erfolgen, langfristig aber zu einer erneuten Erhöhung der Biomasse und des Detritus, wodurch der Bedarf an Sauerstoff exponentiell zunimmt. Wir haben es mit einem typischen Fall einer positiven Rückkopplung zu tun: der Bedarf an Sauerstoff steigert den Bedarf. Das System verbleibt im Ungleichgewicht.
Fazit: Manipulationen haben, wenn überhaupt, nur kurzfristig
Erfolg. Der auch langfristig allein erfolgversprechende Weg ist
eine starke Beschränkung der Nährstoffzufuhr.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de