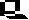
Die Chromosomenanalyse zahlreicher Arten und Gattungen weist ebenso wie die vergleichende Auswertung morphologischer Merkmale auf verwandtschaftliche (phylogenetische) Beziehungen zwischen einzelnen Arten hin. Vielfach sind Genome verwandter Arten so ähnlich (einander weitgehend homolog), daß diploide Primärbastarde entstehen, welche durchaus fortpflanzungsfähig sind:
Es gibt, wie schon erwähnt, zahlreiche weitere Fälle, bei denen Artneubildung auf Addition der Genome beider Elternarten (Alloploidisierung) zurückgeführt werden kann. Andererseits haben gerade die im vergangenen Jahrhundert duchgeführten Experimente von KÖLREUTER, GÄRTNER, NAUDIN, MENDEL u.a. gezeigt, daß man durch Kreuzung zweier Arten nicht ohne weiteres zu neuen stabilen Arten gelangt. Rückkreuzung der Bastarde mit einer oder beiden Elternarten sind oft möglich. Als Folge davon können Genomteile von einer Art in das Genom der anderen überführt werden, deren Genpool bereichern und somit die intraspezifische genetische Variabilität erhöhen. Eine solche zwischenartliche Genwanderung nennt man Introgression. Biologische Arten sind an Standortfaktoren in der Regel gut angepaßt, und nur in den seltensten Fällen ist ihnen ein Bastard hierin überlegen. Meist macht sich Hybridenschwäche und nicht zu unterschätzende Fertilitätsreduktion bemerkbar, so daß die Bastarde den ursprünglichen Arten gegenüber ins Hintertreffen geraten. Vorteile können allerdings bei der Besiedlung neuer, oft "gestörter" Lebensräume entstehen. Dazu einige Beispiele:
Die beiden Arten Geum rivale und Geum urbanum sind in Europa sympatrisch. Die in feuchten Wäldern, in Hecken und vom Menschen geschaffenen Biotopen vorkommende Geum urbanum ist gelb-blühend, die Blüten stehen aufrecht, der Kelch ist zurückgeschlagen, Geum rivale hingegen kommt in Au- und Bruchwäldern sowie auf humösen Wiesen vor. Ihre Blüten sind nickend, die Blütenkronblätter außen rötlich, der Kelch ist anliegend. Geum rivale blüht drei bis vier Wochen früher als Geum urbanum. Geum rivale ist eine typische Bienenpflanze, Geum urbanum mit ihren "offenen", aufrechten Blüten ist weniger spezialisiert; Selbstbestäubung kommt vor. Kommen beide Arten an gleichen Standorten vor, sind Bastatardschwärme nicht selten. Die einzelnen Fl - Bastarde zeigen ein kontinuierliches Spektrum an Übergängen zwischen den beiden Elternarten. Die Mehrzahl der Bastarde ist fertil, in der F2 spaltet sich eine Vielzahl von Kombinationen heraus. Nach diesen Beobachtungen, die auch unter experimentellen Bedingungen reproduzierbar sind (E. MARSTEN-JONES, 1930), müßte man die beiden Arten eigentlich einer Art zurechnen. Geum rivale ist vermutlich einst - geographisch isoliert - in Südosteuropa entstanden. Geum urbanum als weniger spezialisierte Art hat sich in von Menschen geschaffenen Lebensräumen durchgesetzt. Aufgrund ihrer ökologischen Präferenzen bleiben die Elternpopulationen weitgehend stabil, und die Bastarde haben ihnen gegenüber keinen Selektionsvorteil, um sich durchzusetzen. Ein vergleichbares Beispiel stellen auch zwei afrikanische Delphinium-Arten dar.
Adaptive Radiation: Anpassung an neue Bedingungen. Salvia apiana und Salvia mellifera gelten in Südkalifornien als sympatrische Arten mit unterschiedlichen ökologischen Ansprüchen. Die Blüten sind auf Bestäubung durch unterschiedliche Insekten eingerichtet. Salvia apiana wird vornehmlich von großen Holzbienen (Xylocopa) besucht, Salvia mellifera von kleinen bis mittelgroßen Bienen (Osmia, Apis usw.) (K. A. und V. GRANT, 1964). Darüber hinaus wurde eine saisonale Isolation festgestellt. Selten kommt es an natürlichen Standorten zu Hybridisierung; häufig jedoch in neuartigen, durch den Menschen geschaffenen Arealen (C. EPLING, 1947).
Bereits 1935 wies K. M. WIEGAND darauf hin, daß Isolationsbarrieren an natürlichen Standorten funktionieren, in von Menschen geschaffener Umgebung aber zusammenbrechen. Als Beispiel für einen neuen Lebensraum führte er die Bahndämme an. Über weite Entfernungen hinweg verkehrende Züge erwiesen sich als wirkungsvolles Mittel zur Ausbreitung von Pflanzensamen oder anderen Pflanzenteilen. An den neuen Standorten fanden sie Bedingungen, an die sie ggf. nicht adaptiert waren. Ein Ausweg ergab sich durch Hybridisierung mit endemischen, doch verwandten (allopatrischen) Arten.
Anpassung an neue Bestäuber und Lebensräume, Veränderung der Blütenmorphologie: In Kalifornien kommen drei Arten der Gattung Penstemon (Bartfaden, Familie Scrophulariaceae; Abbildungen nordamerikanischer Arten) vor, die sich in Form, Größe und Farbe ihrer Blüten voneinander unterscheiden. Penstemon grinnellii ist in Gebirgskiefernwäldern verbreitet. Die blauen Blütenkronen sind zweilippig und weit geöffnet. Penstemon centrathifolius (Roter Hornist) hat röhrenförmige rote Blüten. Ihr Verbreitungsgebiet erstreckt sich auf die Trockenhänge Südkaliforniens. Die dritte Art (Penstemon spectabilis) findet man in einem relativ neuen, durch Klimaänderung beeinflußten Biotop: auf gestrüppbedeckten Abhängen (Ersatz für vorher dort vorhandene Wälder). Die Blüten sind blau bis schwach violett, sie nehmen in der Form eine Mittelstellung zwischen den beiden erstgenannten ein. Die drei Arten werden normalerweise von folgenden Bestäubern besucht:
Penstemon
grinnellii
von großen Holzbienen (Xylocopa), Penstemon
centranthifolius
von Kolibris, und Penstemon spectabilis von
Wespen und mittelgroßen Bienen.
Aus diesen Befunden konnte die Entstehung der letztgenannten Art wie folgt rekonstruiert werden: Natürliche Bastarde zwischen den beiden ersten Arten (welche immer wieder auftreten) hatten auch in der Vergangenheit zunächst keinen Selektionsvorteil, da die vorhandenen Lebensräume durch die Elternarten belegt waren. Erst nach dem Auftreten buschbestandener Hänge fanden sie eine geeignete ökologische Nische. Wespen und mittelgroße Bienen, die in diesem Lebensraum vorkamen, übernahmen die Bestäuberrolle. Dadurch wurden Blüten bevorzugt, deren Nektar jenen zugänglich war. Es wurden damit solche Genkombinationen selektiert und stabilisiert, die einerseits die Pflanze an eine trockene Umgebung (im Vergleich zu den Wäldern), zum anderen an eine Bestäubung durch Wespen (anstatt durch Kolibris) anpaßten.
Unterschiedlicher Verwandtschaftsgrad zwischen Arten, und die Fähigkeit, Hybride untereinander auszubilden: Die Verträglichkeit der Genome verwandter, teils allopatrischer, teils sympatrischer Arten aus der Compositengattung Laya wurde in den vierziger Jahren von J. CLAUSEN (University of California, Berkeley) analysiert.
Sie können aufgrund der geographischen Verbreitung drei Gruppen zugeordnet werden. In die erste Gruppe gehören zwei Arten (Layia chrysanthemoides und Layia fremantii), von denen die eine im Bereich der San Francisco Bay, die andere östlich davon am Fuße der Sierra Nevada vorkommt. Zur zweiten Gruppe gehören drei Arten (Layia jonesii, Layia munzii und Layia leucopappa), die etwas über 300 km südlich der obengenannten Verbreitungsgebiete zu finden sind. Die dritte Gruppe enthält nur eine, in Südkalifornien beheimatete Art (Layia platyglossa). Ihr Verbreitungsgebiet erstreckt sich aber so weit nach Norden, daß sie sowohl mit den Arten der ersten als auch denen der zweiten Gruppe sympatrisch wird. In jedem Fall besiedelt sie aber andere Biotope als die zuerst genannten Arten. J. CLAUSEN et al. haben unter kontrollierten Bedingungen alle denkbaren Hybridkombinationen hergestellt und die Eigenschaften, vor allem die Fertilität der Nachkommenschaft ermittelt. Alle Arten sind untereinander kreuzbar; die Hybriden zeichnen sich durch üppiges Wachstum aus. Sowohl innerhalb der Gruppe 1 als auch innerhalb der Gruppe 2 beträgt die Fertilität der Hybriden, bezogen auf die Fertilität der jeweiligen Elternarten, 25 bis 30 Prozent. Offensichtlich sind die Isolationsbarrieren zwischen ihnen (noch) nicht genügend gefestigt, so daß man die in Frage kommenden Arten an der Grenze zwischen Art und Unterart (Varietät, Rasse) ansiedeln könnte. Die Fertilität der Hybriden zwischen Arten aus Gruppe 1 und 2 liegt bei nur 5 bis 20 Prozent, d.h., die Gruppen sind untereinander weniger nah verwandt als die Arten innerhalb einer jeden Gruppe. Die isolierte Art aus Gruppe 3 schließlich bildet mit allen übrigen nahezu ausnahmslos sterile Hybriden (0,5-2 Prozent Fertilität). Überall dort, wo sie mit einer der anderen Arten sympatrisch ist, unterbleibt daher nahezu jeglicher Genaustausch.
Morphologische Variabilität und Chromosomenvariation sind nicht immer miteinander korreliert: Clarkia speciosa kommt in Südkalifornien in zahlreichen Varietäten vor, die sich sowohl im Blütenbau als auch z.B. in ihren vegetativen Organen voneinander unterscheiden. Das Verbreitungsgebiet der Unterart Clarkia speciosa nitens liegt im Norden, das von Clarkia speciosa polyantha schließt sich ihm südlich an (H. und M. E. LEWIS, 1955). In einem geographisch kleinen Abschnitt sind beide Arten sympatrisch; Bastardierungen kommen vor. Unabhängig von der Klassifizierung nach morphologischen Kriterien, wurden die Karyotypen der verschiedensten Clarkia speciosa-Populationen untersucht. Alle haben n = 9 Chromosomen. Dennoch lassen sich zwei chromosomale Rassen ausmachen, die durch sieben Translokationen voneinander unterschieden werden können. Während der Meiose der Hybriden entsteht daher ein Ring aus 16 Chromosomen und ein Chromosomenbivalent. Die Situation ähnelt dem, was wir bei Oenothera, Datura und Rhoeo kennengelernt haben. (Anm.: Clarkia gehört wie Oenothera in die Familie der Nachtkerzengewächse: Onagraceae). Die Grenze verläuft quer durch das Verbreitungsgebiet von Clarkia speciosa polyantha, ohne daß sich der Unterschied in irgendeinem ihrer morphologischen Merkmale auswirken würde. Chromosomenvariation und morphologische Variabilität (hier Clarkia speciosa polyantha gegen Clarkia speciosa nitens) erwiesen sich demnach als zwei voneinander unabhängige Größen (W. BLOOM und H. LEWIS, University of California, Los Angeles, 1972).
Alle vorgestellten Beispiele machen deutlich, daß Artbastarde nur dann den Status einer neuen Art gewinnen können, wenn gleichzeitig oder nacheinander mehrere, voneinander unabhängige Umstände eintreten, die die neuen Genotypen stabilisieren und von den Ursprungsarten isolieren. Einen ganz entscheidenden Einfluß scheinen dabei einmal die Umstrukturierung des Genoms, z.B. durch Translokationen, Inversionen, Mutationen usw., zum anderen die Eroberung einer freien ökologischen Nische zu haben.
Wie unsicher einzelne Fortpflanzungsbarrieren sind, und wie flexibel pflanzliche Genome sein können, belegt die Tatsache, daß viele nah verwandte Arten zu Verwandtschaftskreisen zusammengefaßt werden können, innerhalb derer ein Genaustausch möglich ist, wenn die in der Natur wirksamen Barrieren aufgehoben oder gelockert werden. Als ein Maß für die verwandtschaftliche Nähe zwischen zwei Arten (oder Populationen) kann der prozentuale Anteil fertiler Hybriden angesehen werden. Dazu wieder einige Beispiele:
Die Phacelia magellanica-Gruppe (Verbreitungsgebiet: Nordamerika; Familie: Hydrophyllaceae, den Solanaceae verwandt, Abbildungen nordamerikanischer Arten). Dieser Gruppe gehören Arten mehrerer Ploidiestufen an. Von einzelnen Arten kommen diploide, tetraploide und hexaploide Rassen vor. Auf der Stufe der Tetraploiden stehen eine Anzahl von Arten, die untereinander relativ leicht hybridisierbar sind. Es gibt demnach einen wirkungsvolleren Genaustausch zwischen ihnen als zwischen den Ploidierassen der jeweils gleichen Art (R. L. HECKARD, University of California, 1960).
Die Gattung Madia (Verbreitungsgebiet: Nordamerika, Familie: Compositae). Hierzu haben J. CLAUSEN und Mitarbeiter umfangreiche Kreuzungsstudien durchgeführt. Die Ergebnisse sind in der nachfolgenden Abbildung zusammengefaßt. Sie verdeutlichen den unterschiedlichen Verwandtschaftsgrad zwischen den einzelnen Arten und zeigen, was man unter einem "Artkomplex" versteht, in dem es fließende Übergänge im Grad der Hybridisierbarkeit gibt.
Die Gattung Clarkia. Die Ergebnisse der Untersuchungen von H. und M. LEWIS (1955) eignen sich, die phylogenetischen Zusammenhänge zwischen den Arten dieser Gattung darzustellen und die Arten zu einem "Stammbaum" anzuordnen. Strenggenommen handelt es sich hierbei nicht um einen "Baum", denn neben Verzweigungen (Divergenzen) findet man einen hohen Anteil an Konvergenzen (zusammenlaufende Entwicklungen, auf Artbastardierung beruhend). Derartige Verwachsungen (Anastomosen) kommen im Bauplan eines (gesunden !) Baumes normalerweise nicht vor.
Vergleichbare Schemata hat F. EHRENDORFER für die Entstehung der Gattungen aus der Familie der Dipsacaceae sowie für die Achillea millefolium-Gruppe erstellt.
|
|