Seit dem Altertum weiß man, daß physiologische Aktivitäten dem Tagesrhythmus folgen. Die Beobachtungen und Erfahrungen beschränkten sich nicht allein auf Menschen und Tiere, sondern schlossen Aktivitäten von Pflanzen (Heben und Senken von Blättern, Öffnen und Schließen von Blüten) mit ein. Seit dem 18. Jahrhundert wurde die Erscheinung experimentell untersucht. Der Pariser Astronom De MAIRAN erkannte 1729, daß sich die Bewegungen der Pflanzen auch bei ununterbrochener Dunkelheit (Dauerdunkel) fortsetzen, und 30 Jahre später fand der Hamburger J. G. ZINN, daß sich die Blätter der Bohne (Phaseolus coccineus) auch ohne einen Licht-Dunkel-Stimulus heben und senken und daß die Bewegungen weitgehend temperaturunabhängig sind.
C. v. LINNÉ beobachtete, daß das Öffnen und Schließen der Blüten artspezifisch zu bestimmten Tageszeiten erfolgt (Blumenuhr, dargestellt 1755 im Horologium flore confirmandum).

J. v. SACHS (1857, 1863) machte deutlich, daß die Rhythmik durch zwei Komponenten beeinflußt wird, einmal eine erbliche, die festlegt, daß die eigentliche Bewegung rhythmisch abläuft, und zum anderen eine steuernde, die die Phasenlage (den Beginn eines Heben-Senken-Zyklus) festlegt. Seine pflanzenanatomischen Untersuchungen der Blattstielbasis von Oxalis carnea gaben Aufschluß über die Struktur des Gelenks und bildeten die Grundlage, um das Heben und Senken der Blätter als eine Turgorbewegung zu charakterisieren. Im Gegensatz hierzu sind die Bewegungen der Blütenblätter Wachstumsbewegungen.
 Die Bedeutung der Autonomie der Rhythmik wurde zu Beginn unseres
Jahrhunderts durch R. SEMON, und später auch durch W. PFEFFER
(ab 1907) erkannt. Den endgültigen Beweis erbrachten E. BÜNNING
und K. STERN (seinerzeit, 1930, am Botanischen Institut der Universität
Jena), indem sie die Reaktionen von Phaseolus multiflorus
und anderen Arten unter thermokonstanten Laborbedingungen nach
einem vorgegebenen Lichtprogramm (Hell-Dunkel-Wechsel) analysierten.
Durch diesen experimentellen Ansatz wurde sichergestellt, daß
es sich um autonome Bewegungen, also um eine endogene Rhythmik
handelt, die durch den Außenfaktor Licht gesteuert werden
kann. Bei Wegfall der Steuerung weicht die Periodenlänge
signifikant (art- und individuenspezifisch) von 24 Stunden ab,
so daß man von einer circadianen Rhythmik sprechen muß
(E. BÜNNING, 1986).
Die Bedeutung der Autonomie der Rhythmik wurde zu Beginn unseres
Jahrhunderts durch R. SEMON, und später auch durch W. PFEFFER
(ab 1907) erkannt. Den endgültigen Beweis erbrachten E. BÜNNING
und K. STERN (seinerzeit, 1930, am Botanischen Institut der Universität
Jena), indem sie die Reaktionen von Phaseolus multiflorus
und anderen Arten unter thermokonstanten Laborbedingungen nach
einem vorgegebenen Lichtprogramm (Hell-Dunkel-Wechsel) analysierten.
Durch diesen experimentellen Ansatz wurde sichergestellt, daß
es sich um autonome Bewegungen, also um eine endogene Rhythmik
handelt, die durch den Außenfaktor Licht gesteuert werden
kann. Bei Wegfall der Steuerung weicht die Periodenlänge
signifikant (art- und individuenspezifisch) von 24 Stunden ab,
so daß man von einer circadianen Rhythmik sprechen muß
(E. BÜNNING, 1986).
Bild links: Professor Erwin BÜNNING unterwegs mit dem Nordpilen zur jährlichen Lappland-Exkursion (1963), die er für Studierende der Biologie an der der Universität Tübingen organisierte.

Phaseolus coccineus. Typischer Verlauf tagesperiodischer Blattbewegungen unter Dauerlichtbedingungen (schwache Lichtintensität). Innerhalb von sechs Tagen erfolgt gegenüber dem normalen Tag eine Phasenverschiebung um ungefähr 17 Stunden; die Periodenlänge beträgt also etwa 27 Stunden. (Kreisbögen in 24-Stunden-Abständen). (E. BÜNNING und M. TAZAWA, 1957).
Innerhalb der Art Phaseolus coccineus wurden Pflanzen mit einem Zyklus von 23 und andere mit einem von 26 Stunden identifiziert. BÜNNING fand, daß die Periodenlänge über mehrere Jahre und Generationen hinweg konstant blieb. Dazu wurden die einzelnen Pflanzen fortlaufend geselbstet und deren Nachkommenschaften analysiert. In einer parallel angesetzten Versuchsserie wurden 23-Std.-Pflanzen mit 26-Std.-Pflanzen gekreuzt. Die Periodenlänge bei den`Nachkommen in der Fl bildete das Mittel zwischen beiden Elternstämmen. In späteren Generationen kamen die Periodenlängen der Eltern wieder zum Vorschein.
In neuerer Zeit werden Untersuchungen zur endogenen Rhythmik in zunehmendem Maße an einzelligen Organismen durchgeführt. Als Maß der Rhythmik werden nicht Bewegungen, sondern Stoffwechselaktivitäten (z.B.Sauerstoff-Produktion, bedingt durch die Photosynthese) herangezogen. So konnte V. G. BRUCE 1972 durch Kreuzung zweier Chlamydomonas-Stämme (einer mit einer 24-Std.-, der andere mit einer 26-Std.-Periode) belegen, daß die Phasenlänge durch die Aktivität eines einzigen Gens verursacht werden kann.
Durch Isolation und Einsatz weiterer Chlamydomonas-Stämme wurde eine additive Wirkung periodenverlängernder Gene nachgewiesen (BRUCE, 1974): Wenn eine Mutante die Periode um n, eine andere um m Stunden verlängert, zeigt die durch Kreuzung erhaltene Doppelmutante eine Verlängerung um n + m. Wie an diesem Beispiel gezeigt, kommt eine endogene Rhythmik nicht nur in Bewegungen, sondern auch im Wechsel von Stoffwechselraten und anderen Aktivitäten zum Ausdruck. Da der Tag-Nacht-Rhythmus zu den wenigen konstanten Parametern unserer Umwelt gehört, war es zwingend, daß sich im Verlauf der Evolution der Tiere und Pflanzen Mechanismen entwickeln mußten, um Zeit zu messen und auf den Tag-Nacht-Wechsel zweckentsprechend zu reagieren. Auffallenderweise sind diese Erscheinungen niemals bei Bakterien beobachtet worden. Das ist verständlich, denn ihre Generationsdauer beträgt unter günstigen Bedingungen, je nach Art, Minuten oder Stunden; die Tagesperiode tritt im Lebenszyklus einer Bakterienzelle gar nicht in Erscheinung.
Zu den periodischen Prozessen (der Pflanzen) gehören u.a. auch die Entleerung der Sporangien bei manchen Algen (und Pilzen) und Aktivitäten, die sich auf der Transkriptions- und Translationsebene sowie der Regulation des Stoffwechsels manifestieren.
Normalerweise laufen die verschiedensten circadiaen Vorgänge in der Zelle immer in fester Phasenbeziehung zueinander ab. Wird die Phasenlage einer Reaktion im Experiment durch äußere Faktoren verschoben, ändert sich die Phasenlage aller mit ihr in Beziehung stehenden Reaktionen um den gleichen Zeitabschnitt. Der Effekt kann auch durch Mutation hervorgerufen werden, da dabei sich ebenfalls alle an einem betreffenden Objekt untersuchten circadianen Funktionen in gleichartiger Weise ändern. Rein formal können wir uns das an einem einfachen Modell veranschaulichen. Es beruht auf einer Zusammenschaltung mehrerer Regelkreise (A, B, C, D...), die in der Abbildung durch Kreise, oder besser gesagt, durch gleich große Räder symbolisiert sind, und zeigt, daß die Phasen der einzelnen Regelkreise gegeneinander versetzt sind, somit beliebige Werte annehmen können. Nunmehr stellt sich für uns die Frage: Wie messen die Organismen die Zeit und wie werden die einzelnen Aktivitäten synchronisiert?
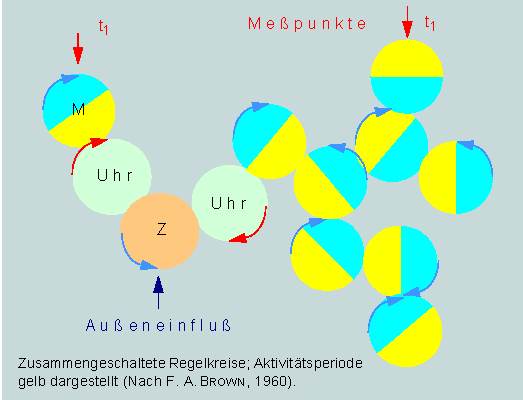
Es gibt gut begründete Hinweise darauf, daß Zeitmessungen mit Hilfe von Schwingungen eines "Oszillators", und nicht etwa nach dem Sanduhrprinzip erfolgen. Damit liegt die Analogie zu einer Taschenuhr auf der Hand. Bei ihr wird die Energie einer Feder auf eine Unruhe (einen Oszillator mit einer Schwingungsdauer im Sekundenbereich) und von dort über einen ungleichen Übersetzungsmechanismus (verschieden große Rädchen) auf den großen und gleichzeitig auf den kleinen Zeiger übertragen, steuert dadurch synchron eine l -Std.- und eine 12 -Std.-Periode.
Alle komplizierten funktionellen Strukturen, ganz gleich, ob Organismen oder technische Geräte, sind als geregelte Systeme zu verstehen, in denen eine Vielzahl von Regelkreisen miteinander vernetzt sind. Aus der Theorie eines Regelkreises heraus ergibt sich, daß der Istwert (= der zu einem bestimmten Zeitpunkt tatsächlich gemessene Wert) stets um den Sollwert oszilliert. Daraus folgt zwangsläufig, daß in jedem Organismus a priori Schwingungen mit den verschiedensten Frequenzen, also mit Perioden von Bruchteilen einer Sekunde, von mehreren Sekunden, Minuten, Stunden usw. auftreten. Zur Anpassung an seine Umwelt selektiert der Organismus offenbar aus den verfügbaren Regelkreisen (Reaktionen) diejenigen mit der optimalen Frequenz. Zur Ergänzung sei vermerkt, daß man außer einer Tagesrhythmik eine Jahresrhythmik (s. hierzu u.a. Photoperiodismus) und z.B. bei einigen Meeresalgen (und -tieren) eine lunare Rhythmik (messbar als tidenabhängige Rhythmik) kennt.
Bei der Besprechung des Primärstoffwechsels haben wir bereits eine Anzahl von Regelmechanismen kennengelernt. Zu den rasch ansprechenden gehört die sogenannte Endprodukthemmung, durch die die Konzentrationen bestimmter Metaboliten gesteuert werden. Die Periodenlänge liegt in der Größenordnung von Sekunden/Minuten. Im Gegensatz hierzu wird die Regulation über die Genexpression in Zeiträumen von Stunden bewerkstelligt.
So nützlich die vorliegenden Ergebnisse und Modelle zum prinzipiellen Verständnis biologischer Uhren auch sein mögen, so wenig haben sie zur Aufklärung des tatsächlichen Mechanismus der circadianen Rhythmik beigetragen. An der Regulation des Stoffwechsels sind Enzyme beteiligt, und deren Aktivität ist streng temperaturabhängig. Es gilt die Faustregel, daß sich die Aktivität bei einer Temperaturerhöhung um 10 Grad C im physiologischen Temperaturbereich verdoppelt. Man sagt daher, der Q10-Wert sei 2. Für tagesperiodische Aktivitäten gilt jedoch über einen weiten Temperaturbereich hinweg ein Q10 von 1, und das wiederum heißt, daß die rhythmischen Erscheinungen weitgehend temperaturunabhängig sind und deshalb nicht durch wechselnde Enzymaktivitäten erklärt werden können. Es ist bisher auch von keinem Enzym erwiesen, daß es zum "Uhrwerk" gehört. Daher muß mit der Möglichkeit gerechnet werden, daß alle gefundenen Enzymrhythmen nur periphere, d.h., von der Uhr gesteuerte Rhythmen zum Ausdruck bringen. Ein abrupter Temperaturwechsel (Unterkühlung) bringt jedoch, wenn er zum richtigen Zeitpunkt erfolgt, die Rhythmik aus dem Gleichgewicht. Die Analyse der einzelnen Phasen auf ihr Verhalten gegenüber Temperaturerniedrigung ergab, daß sie in einigen Abschnitten des Zyklus keine oder kaum eine Wirkung zeigte, in anderen dagegen eine starke Verzögerung hervorrief, d.h. also, daß die Uhr in den verschiedenen Phasen unterschiedlich stark auf äußere Reize reagiert.
In gewisser Hinsicht vergleichbare Ergebnisse erhält man durch Applikation kurzer Lichtblitze (Störlicht: 1-2 min. pro Tag). Auch dabei hängt es davon ab, in welcher Phase die Pflanzen den Lichtreiz erhalten. Empfindliche Phasen werden als photophil, unempfindliche als skotophil (photophob) bezeichnet.
Bislang haben wir die Lichtqualität außer acht gelassen. Über weite Spektralbereiche bleibt die Wirkung gleich, eine Ausnahme bildet der Rotlichtbereich:
Wie an anderer Stelle dargelegt , gehört das Phytochromsystem zu den wichtigsten Lichtrezeptoren der Pflanzen. Es wird durch dunkelrotes Licht inaktiviert, durch hellrotes aktiviert. Das Verhältnis beider Lichtqualitäten zueinander determiniert die Menge an aktivem Phytochrom. J. v. SACHS hatte schon im letzten Jahrhundert postuliert, daß nicht das Licht an sich, sondern Änderungen der Lichtintensität als auslösender Reiz (Zeitgeber) der circadianen Rhythmik anzusehen seien. Mit der Änderung der Tageslichtintensität (morgens und abends) läuft auch eine Änderung der spektralen Zusammensetzung parallel. Tagesrestlicht enthält nämlich im Vergleich zum Tageslicht einen relativ hohen Dunkelrotanteil. Dunkelrotlicht verkürzt nicht nur die Periodenlängen, sondern führt zu einem schnellen Abklingen der Rhythmik. Wenn nun ein Faktor gleichzeitig kurze Perioden verursacht und die Rhythmik rasch ausklingen läßt, liegt der Schluß nahe, daß er einen Schwingungsabschnitt vorzeitig abbrechen läßt. Daraus resultiert, daß der gegenläufige Schwingungsabschnitt zwangsläufig kürzer ist. Aufeinanderfolgende Wiederholungen führen daher zu einer Dämpfung, und schließlich zu einem Ausklingen der Schwingung.
Ein weiteres Problem wäre die Frage nach dem Ort der Lichtperzeption. Wie bei anderen lichtinduzierten Reaktionen wird auch hier bei höheren Pflanzen zwischen Zellen, die den Reiz empfangen, und solchen die die rhythmische Aktivität ausüben, unterschieden. Für mehrere Arten ließ sich nachweisen, daß die Lichtabsorption primär in den Epidermiszellen der Laubblätter erfolgt. Oft enthalten sie speziell geformte Zellen mit Linsenwirkung (Ozellen), die auf Lichtabsorption spezialisiert sind. Ebenso mag das Fehlen von Chlorophyll in der Epidermis der Blattoberseite vorteilhaft sein, denn es wird als Konkurrent der Lichtabsorption ausgeschaltet, womit ein Ansprechen auf geringe Lichtmengen (Größenordnung 1 Lux) erreicht wird.
Das erwähnte Phytochromsystem ist sicherlich nicht der alleinige Lichtrezeptor. Bei vielen Pflanzen ist Blaulicht stärker, teilweise sogar ausschließlich wirksam. Derartige Unterschiede verdeutlichen, daß die absorbierenden Pigmente nicht Bestandteile der Uhr, wohl aber an sie gekoppelt sind.
Abschließend: Was kann man über den Oszillator selbst aussagen? Es bereitet kaum Schwierigkeiten, den Zellkern der einzelligen Riesenalge Acetabularia zu entfernen. Kernlose Stücke bleiben über Wochen am Leben und zeigen deutliche tagesperiodische Rhythmen (z.B. in ihrer Photosyntheseaktivität). Daraus folgt, daß der Kern zur Aufrechterhaltung der Rhythmik nicht benötigt wird. Implantiert man jedoch kernfreien Fragmenten einen Kern aus einer Zelle, die auf inverse Rhythmik programmiert war, übernimmt das Fragment die Phasenlage des Kerns. Damit können ihm zweifelsohne Zeitgeberfunktionen zugeschrieben werden. Nun lassen sich mit dem Acetabularia-System noch eine Reihe weiterer Experimente anstellen. Aus der Molekularbiologie weiß man, daß es eine Anzahl spezifischer Inhibitoren für die einzelnen Teilschritte der Genexpression gibt. So ließ sich zeigen, daß die Applikation von Chloramphenicol und Rifampicin die Rhythmik nicht beeinflußt, während die von Puromycin oder Cycloheximid schwere Störungen der Uhr verursacht. Rifampicin hemmt die Transkription (inhibiert die DNS-abhängige RNS-Polymerase), Chloramphenicol hemmt die Translation an Ribosomen der Prokaryoten sowie der Ribosomen in Chloroplasten und Mitochondrien, und Cycloheximid schließlich hemmt spezifisch die Proteinbiosynthese an cytoplasmatischen Ribosomen, Puromycin unterbindet jede Art der Proteinsynthese. Aus den genannten Inhibitionsexperimenten folgt, daß die circadiane Rhythmik nicht durch die Transkription und nicht durch die Translation in Chloroplasten oder Mitochondrien, wohl aber durch die an cytoplasmatischen Ribosomen beeinflußt wird. Weitere Experimente führten zu der Annahme, daß die Synthese eines membrangebundenen Proteins, das seinerseits seine Eigensynthese blockiert, an der Initiation der Rhythmik mitwirkt und als ein Kandidat für den Oszillator in Frage kommt (H. G. SCHWEIGER, 1982). Auch hier bedarf es noch zahlreicher Versuche, um die Annahme abzusichern und ihre Gültigkeit für andere Systeme zu verifizieren.
|
|